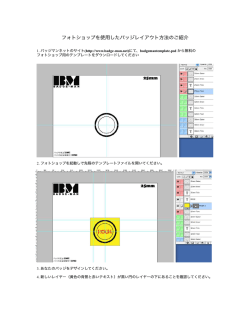
日薬理誌(Fo伽織α""αcol.〃":.)140.19∼23(2012) ; 玲 言 二 I シナプス入力のリアルタイムイメージング 小林千晃,高橋直矢,池谷裕二 要約:脳は多様な情報を柔軟に処理することで多彩な 不十分である. 機能を発揮している.これまでに,個体レベルから単 とともに,従来のアプローチだけでは脳の多様性を説 今回,我々は新たに開発した大規模な画像法によっ て,単一ニューロンのシナプス活動を多数のスパイン (注:興奮性シナプスの後部に存在する突起構造)か ら同時に記録することに成功した.その結果,神経回 路がシナプスレベルで精密に編み込まれていることを 明することは不十分であることもわかってきた.神経 明らかにし(1).脳機能の多様性の根源に迫ることが 回路を構成するニューロンは,互いにシナプスを介し できた.本稿では,樹状突起によるシナプス入力の統 て結合することで情報を伝達している.近年,樹状突 合処理能力について,我々の発見に至るこれまでの研 起に存在する多数のシナプスを通じて,単一のニュー ロンでも高度な情報処理を行うことが可能であること 究の歴史について説明するとともに,今回我々の開発 した大規模スパインカルシウム画像法の応用の可能性 が示唆されている.たとえば,ニューロンは,シナプ に触れる. 一細胞レベルに至る多くの研究がなされ,脳の機能の 多様性がどう生じているのかという疑問が徐々に解消 されてきた.しかし,こうした研究成果が蓄積される ス活動の時空間パターンを非線形に積算することで, 情報のフィルタリングや論理演算を行なっている.ま 1.樹状突起スパイン た,シナプス可塑性を通じて積極的に神経回路網を書 ニューロンには通常,軸索と樹状突起という2種類 き換えることで,より効率的に情報の多様性と安定性 を実現している.したがって,シナプス活動を詳細に の神経線維が存在している.樹状突起上には,単一ニ ューロンあたり数千から数万に及ぶスパインと呼ばれ 観察することは,ニューロンの作動原理を解明するう る後シナプス構造が存在し,上流のニューロンの活動 えで必須である.今回,我々は,機能的スパインカル をシナプス入力として受け取っている.スパインは シウム画像法の開発により,ニューロンの‘情報処理を 100年以上前から存在が知られていおり(2),その多様 シナプスレベルで解明することができるようになった. な形態も明らかとなっていた(図1).しかし,スパイ 本稿では,同実験手技の開発に至る経緯とともに,今 ンは直径が1マイクロメーターにも満たない微細な突 起構造であり,電気生理学のように記録電極を刺入し 後の展望についてもふれる. はじめに てそこでのシナプス活動を記録することはきわめて困 した脳の機能を十分には説明できていないまた多く 難である.近年,そうした実験的な制約を解消する手 段として,神経活動への光学的なアプローチが注目を 浴びている.二光子レーザー顕微鏡やカルシウム蛍光 プローブ,電位感受性色素が開発・改良され,スパイ ンの形態とその機能的役割が序々に明らかとなってき の難治性中枢神経系疾患において,シナプスに器質的 ている(3). 脳は多様な情報を精密に処理するとともに,多様な 機能を発揮できる.しかし,従来行われてきた個体レ ベル,神経回路レベル,細胞レベルの研究では,こう な異常が明らかになっているにも関わらず,その構造 が微小であるため,機能異常や治療法の解明はいまだ キーワード:ニューロン,シナプス,樹状突起,スパイン,イメージング 東京大学大学院薬学系研究科薬品作用学教室(〒113-0033東京都文京区本郷7-3-1) E-mail:ikegaya@mol.f.u-tokyo・ac.jp原稿受領日:2012年4月20日,依頼原稿 T i t l e : R e a l t i m e i m a g i n g o f s y n a p t i c i n p u t s . A u t h o r : C h i a k i K o b a y a s h i , N a o y a T a k a h a s h i , Y u j i I k e g a y a 20 小林千晃・高橋直矢,池谷裕二 ことが報告されている(7,8).すなわち,シナプス入 力の時空間構造そのものがその統合過程,さらにはニ ューロンの出力発火を決定する重要な要因となりうる ( 9 ) . 3.従来の樹状突起研究 ニューロンが,より効率的に活動電位を発生させる には,樹状突起が電位感受性のチャネルや受容体を介 して,シナプス入力を超加算的に統合することが必要 である(10).しかしながら,樹状突起スパイクなどの, 樹状突起がもつシナプス入力の統合機能に関する研究 の多くは,樹状突起を電気的に刺激することや,グル タミン酸アンケージングなどを用いた人工的なシナプ 図1錐体細胞樹状突起の蛍光画像 錐体細胞樹状突起には複数のスパインが存在する.様々な形態の スパイン(矢印)が存在することがわかる. ス入力を惹起することで行われることが一般的であっ た.そのため,ニューロンが潜在的にシナプス入力を 多様に処理する能力をもつことは示されてきたものの, 2.シナプス入力を統合する場としての樹状突起 シナプス入力は樹状突起において統合され,細胞体 内在する神経活動の中でニューロンが実際にどのよう な時空間特性のシナプス入力を受容しているのかにつ いてはほとんどわかっていない. へと伝播される.近年,入力の統合過程において,樹 状突起が動的な特性を示すことが明らかとなってきて ー入力と分散入力の二つのモデルが想定される(図2). いる.樹状突起を静的な伝導体ケーブルと捉えた場合, クラスター入力は,上述した樹状突起スパイクを発生 シナプス入力は,興奮性シナプス入力と抑制性シナプ させる条件を満たしており,非線形演算が可能である. ス入力の総和として樹状突起上で加算され,ケーブル その一方で,分散入力では,樹状突起スパイクの発生 特性により指数関数的に減衰しながら,細胞体へ届く と考えられる.しかしながら,樹状突起には,電位感 受性NaチャネルやCaチャネル,NMDA受容体など の電位感受性のコンダクタンスが分布しており,細胞 体に届く電気信号は,興奮性シナプス入力と抑制性シ ナプス入力の単純な総和とはならない(4).たとえ同 じ入力数であっても,時間や空間的に異なる複数のシ ナプス入力は,異なる加算過程を経て,異なる電位変 は期待できない.本来,樹状突起の非線形演算は,こ 化を生じる(5). さらに高度な相互作用の例は,樹状突起スパイクで シナプス入力の時空間構造を考える上で,クラスタ うした実際のニューロンで生じうるシナプス入力の時 空間構造に関して議論されるべきであるはずが,技術 的な制約によりアプローチできず,シナプス入力の時 空間特性について明確な解答は得られていなかった. 4.シナプス活動イメージング 生理活動下におけるシナプス入力の時空間特性につ いてはほとんどわかっていない(11).この主たる理由 は,これまでの実験手法の限界に因るところが大きい. ある(6).樹状突起スパイクは,樹状突起の限局した 従来,ニューロンのシナプス活動を記録する際には, 領域が,集中してシナプス入力を受けることで,電位 細胞体からのパッチクランプ記録が汎用されてきた. 感受性チャネル等が活性化して局所で生じる活動電位 同手法では細胞体にガラス電極を刺入し,シナプス活 である.樹状突起スパイクによって入力が超加算的と 動を電気信号として記録する.しかしながら,この場 なるため,通常のシナプス入力と比べ,より効率的に 合,シナプス活動の時系列パターンを計測できるもの 細胞体を脱分極させることができる. の,どの樹状突起,あるいはどのスパインが入力を受 こうした複雑な加算機構の存在により,ニューロン けたのかを知ることは不可能である.つまり,電気生 は非線形演算子として働くことができる.また,グル 理学的な記録では空間解像度が不足している.そこで, タミン酸アンケージング法や電気生理学的手法を用い シナプス入力の空間構造を知るために,多数のスパイ て人工的にシナプス入力を惹起した場合,シナプス入 ンより同時に活動を計測する画像法開発が求められて 力の加算様式が,樹状突起の形態,各種イオンチャネ いた. ルおよび受容体の発現パターンにより様々に変化する シナプス入力時には,スパインにおいてNMDA受 ■■■総説 シナプス入力のリアルタイムイメージング クラスター入力モデル 分散入力モデル 上遼ニューロン集団1 上流ニューロン築団1 ン ン 上流 21 上滝 図2こつのシナプス入力モデル プレシナプス細胞集団からの入力が,特定の樹状突起に集中するクラスター入力モデル(左)か.樹状突起全体に分散する分散入力モデル(右) の二つの可能性がある.クラスター入力モデルではシナプス入力が集中するため.樹状突起スパイクを起こすことが可能である.(文献1より許 可を得て改変) 容体を介して一過的にカルシウムが流入するため,カ に異なる3つの特長を持つ.すなわち,①広域の視野 ルシウム蛍光指示薬によって可視化しうることが, から高速で映像を連続取得できる,②退色が遅い,③ Yusteらによって1995年に示された(12).したがって, 光毒性が低い,である.このような特長は,スパイン この蛍光変化を一斉に画像化することによって,シナ のように微細な構造物から長時間安定してイメージン プス入力の時空間構造を明らかにできるはずである. グを行うのに適していると考えた.まず我々は.CAS スパインは微小な構造であるために,シナプス活動に 錐体細胞からホールセル記録を行い,カルシウム蛍光 応じた蛍光指示薬の蛍光強度変化が微量である.従来 指示薬を導入し(図3a),同顕微鏡を用いてカルシウ はこれを検出することは困難であったが,同年に ム活動を記録したところ,広い視野から高い時空間解 CCD(chargecoupleddevice)カメラと優れたカルシ 像度でイメージングできることがわかった(図3c.d) ウム蛍光センサーを用いることで,十分なSN比をシ (15).そして,さらに実験条件の最適化を図ることで, グナルとして捉えることができることが報告された 100個以上のスパインから同時にシナプス活動を観察 (13).近年では共焦点レーザー顕微鏡の技術が発達し, することに成功した(図3e).また,イメージング後 スライス標本の深部に励起レーザーを散乱を防ぎつつ に樹状突起の形態を3次元的に構築することで,記録 照射し,様々な領域からスパインを可視化できるよう されたスパインの空間配置を正確に把握することがで になった.そうした技術の進歩の一方で,レーザーに きる(図3b).これにより,「どの」スパインが,「い よる光毒性や色素の退色の問題が新たに浮き彫りにな つ」,「どこで」入力を受けたのかという,シナプス活 った.この問題を解消したのが,二光子レーザー顕微 動の時空間特性を捉えることが可能になった(図3). 鏡である(3).二光子レーザー顕微鏡は,その特徴に より,光毒性と色素の退色を抑え,標本を非侵襲的に 観察できるとともに,高い空間解像度を有する.また, 5.クラスター入力 樹状突起が動的な演算子として働くには,樹状突起 共焦点顕微鏡よりもさらに深部にまでレーザーが届く スパイクなどのように,入力を非線形に統合する必要 という利点を併せ持つため,invivoにおけるシナプス がある(5,16).一方,樹状突起スパイクを発生させる 活動を捉えることもできるようになった(14).ただ, には,これまで述べてきたように,樹状突起のある特 こうした手法で得られるスパインの活動情報は,せい 定の領域に対し,まとまった入力(クラスター入力) ぜい数個程度のスパインであり,さらに時間分解能も が必要である(図2)(17).近年,樹状突起の演算能力 十分ではないため,シナプス入力の時空間構造を明ら に関して多く研究がなされているが,クラスター入力 かにすることは不可能であった. が実際の神経回路で生じているのかという疑問の解決 では,どうすればシナプス入力の時空間特性を観察 には至っていない. することができるだろうか.我々はニポウ板型共焦点 シナプス入力がクラスター化している可能性につい レーザー顕微鏡の特徴に着目した(15).この顕微鏡は ては,可塑性の研究からも支持される(10,18).シナ 微弱なレーザーを多数用いることで高速に映像を得る プス可塑性の代表的な型であるLTP(longterm 仕組みで,従来型の共焦点レーザー顕微鏡とは本質的 potentiation)は,シナプス結合強度を増強し,シナプ 22 小林千晃,高橋直矢,池谷裕二 C OS 0.3s ARF ” y i 2低m dスパイン#29 3s 50% 4 " 1 0 110 1 e” 共焦点面 転入やて× 1 0 20 40 60 80 100 120(s) 図3大規模スパインイメージング法によるシナプス活動の観察 (a)イメージングされたCA3錐体細胞.パッチクランプ記録によりカルシウム蛍光指示薬Fluo-5Fを負荷した.(b)三次元橘築された樹状突 起形態.aの四角枠の部位に相当する.(c)スパインの自発劇なカルシウム活動の時系列変化bの白枠の部位に相当する.(d)スパインのカ ルシウム蛍光トレース.(e)bに示す137個のスパインの活動パターンを表すラスタープロット.各点が個々のカルシウムイベントを示す. スを介した情報伝達が効率よく行えるよう変更する機 情報伝達を行っているかどうかの決定的な証拠はいま 構である.近年,ロPを示したスパインに近接したス だ得られていない. パインで皿Pがより誘導されやすくなることが示さ れている.ある特定のスパインで皿Pが誘導されると, 6.大規模スパインカルシウム画像法 タグタンパク質によって付近のスパインがラベル化さ 上記で述べたように,近接したスパインは局所的な れる.そのタグタンパク質の存在が,周囲のスパイン 可塑性機構を通じてクラスターを形成し,シナプス入 のロP誘導を容易にすると考えられる(18).また, 力は樹状突起上で時空間的に集中すると考えられる. GTPaseの一種であるRasは,LTPによって特定のス その結果,樹状突起スパイクに代表される超加算的な パインで活性化されると,周囲のスパインヘと拡散し, シナプス入力の演算が可能になると想定される(図2). 皿Pが誘導されやすくなることもわかっている(19). 今回,我々は大規模スパインカルシウム画像法を用い さらに,樹状突起スパイクの一種であるNMDA受容 ることで,同期したシナプス入力が樹状突起上の特定 体を介したNMDAスパイクは,カルシウム流入を伴 の領域のスパインに集中することを明らかにした(図 いながら局所的に樹状突起を伝播する.その結果, 4a)(1).クラスター入力のサイズは8マイクロメート NMDAスパイクの発生した特定の樹状突起において, ル以内で統計学的に有意であった.クラスター入力は カルシウム濃度がより上昇し,LTP誘導シグナルは賦 電気刺激による人工的なシナプス入力の惹起では生じ 活化されやすくなる(10).以上の機構により,皿Pは なかった(図4b).さらに,NMDA受容体を慢性的に 空間的に近接したスパイン集団で発生することが想定 遮断した神経回路(図4c)や,未成熟な神経回路(図 される.これらの研究から,樹状突起の局所で皿Pが 4d)においても観察されなかったことから,シナプス 発生し,スパインのクラスター化を加速しているとい 入力のクラスター化はNMDA受容体に依存した神経 う仮説が,いよいよ現実味を帯びてきたわけである 回路網の発達によって獲得されるものと考えられる. (図2).だが,ニューロンがクラスター入力を介した 最近,invivoで運動学習によって特定のスパインが ■■■総説 23 シナプス入力のリアルタイムイメージング 鰭発活動 30 重︶掛漣 840 a︵誤﹀掛鯉 b 01020304050600102030405060 同期したスパイン間の距離(Mm) 同期したスパイン間の距離(urn) ︵誤︶掛鯉 0 1 0 2 0 3 0 4 0 5 0 同期したスパイン間の距離(urn) 840 ︵ま︶掛鯉 840 C 0 1 0 2 0 3 0 4 0 5 0 同期したスパイン間の距離(urn) 図4シナプス入力のクラスター化 スパインがあるスパインと1叩ms以内に同期入力を受ける確率を縦軸に.同期入力を受けたスパイン間の距離を横軸に示した.(a)自発活動 下におけるシナプス入力.(b)放線状層を刺激によって誘発したシナプス入力.チヤンスレベルを灰色線で、その95%信頼区間を灰色域で示し た.自発活動下では距離の近いスパイン同士が同期しやすいことがわかる.(c)NMDA受容体阻害薬存在下で.組織培養した場合,シナプス入 力はクラスター化しない.(d)未成熟回路では,シナプス入力のクラスター化は見られない.(文献1より許可を得て改変) 反復的に活性化されると,そのスパインを中心とした か.シナプス画像法の医薬分野への応用は大きな可能 局所に新たなスパインが形成されるという研究結果が ‘性を秘めている. 発表されている(20).我々の発見に加えて,こうした 文 献 発見は,樹状突起の非線形演算を行うとする仮説とよ く一致している. 1 ) T a k a h a s h i N , e t a l . S c i e n c e . 2 0 1 2 : 3 3 5 : 3 5 3 3 5 6 . 2)YusteR.Neuron、2011:71:772-781. 7.大規模スパインカルシウム画像法の応用 3 ) D e n k W , e t a l . C u r O p i n N e u r o b i o l . 1 9 9 6 ; 6 : 3 7 2 3 7 8 . 4)TakahashiN,etal.ProcNatlAcadSciUSA.2010;107:1024410249. 多くの難治性中枢神経系疾患において,シナプスに 器質的な異常が認められている.たとえば,アルツハ イマー病,ダウン症,統合失調症などの中枢神経系の 機能障害に起因する病態において,シナプス数の減少 が報告されている(21-23).こうした変化はシナプス 活動の時空間パターンにも大きく影響すると推測され る.各種の神経疾患モデルを対象にスパイン画像法を 用いれば,シナプス活動レベルでの病理解析が可能で ある.また神経疾患に対する既存薬の中には,神経伝 達物質量の調節などシナプス機能に作用すると期待さ れるものが多いこれら薬物のシナプス活動に与える 影響を精査すれば,薬の有効性を再評価でき,新たな 創薬ターゲットの開拓にもつながるのではないだろう 5 ) H a u s s e r M , e t a l . C u r O p i n N e u r o b i o l . 2 0 0 3 ; 1 3 : 3 7 2 3 8 3 . 6 ) L a r k u m M E , e t a l . S c i e n c e . 2 0 0 9 ; 3 2 5 : 7 5 6 7 6 0 . 7 ) H a u s s e r M , e t a l . S c i e n c e . 2 0 0 0 ; 2 9 0 : 7 3 9 7 4 4 . 8 ) L o n d o n M , e t a l . A n n u R e v N e u r o s c i . 2 0 0 5 ; 2 8 : 5 0 3 5 3 2 . 9 ) P o i r a z i P , e t a l . N e u r o n . 2 0 0 3 ; 3 7 : 9 8 9 9 9 9 . 1 0 ) L a r k u m M E , e t a l . C u r O p i n N e u r o b i o l . 2 0 0 8 ; 1 8 : 3 2 1 3 3 1 . 1 1 ) J i a H , e t a l . N a t u r e 、 2 0 1 0 ; 4 6 4 : 1 3 0 7 1 3 1 2 . 1 2 ) Y u s t e R , e t a l . N a t u r e . 1 9 9 5 ; 3 7 5 : 6 8 2 6 8 4 . 1 3 ) S e g a l M . J N e u r o s c i M e t h o d s . 1 9 9 5 ; 5 9 : 2 5 2 9 . 1 4 ) S v o b o d a K , e t a l . S c i e n c e . 1 9 9 6 ; 2 7 2 : 7 1 6 7 1 9 . 1 5 ) T a k a h a s h i N , e t a l . N e u r o s c i R e s 、 2 0 0 7 ; 5 8 : 2 1 9 2 2 5 . 1 6 ) M a g e e J C . N a t R e v N e u r o s c i . 2 0 0 0 ; 1 : 1 8 1 1 9 0 . 17)SilverRA.NatRevNeurosci.2010;11:474-489. 1 8 ) G o v i n d a r a i a n A , e t a l . N a t R e v N e u r o s c i . 2 0 0 6 ; 7 : 5 7 5 5 8 3 . 1 9 ) H a r v e y C D , e t a l . S c i e n c e . 2 0 0 8 ; 3 2 1 : 1 3 6 1 4 0 . 2 0 ) M i n F , e t a l . N a t u r e . 2 0 1 2 ; 4 8 3 : 9 2 9 5 . 2 1 ) S t e p h e n W S , e t a l . N e u r o b i o l A g i n g 、 2 0 0 6 ; 2 7 : 1 3 7 2 1 3 8 4 . 2 2 ) L e i s A , e t a l . A r c h G e n P s y c h i a t r y . 2 0 0 0 ; 5 7 : 6 5 7 3 . 2 3 ) F e r r e r I , e t a l . A c t a N e u r o p a t h o l . 1 9 9 0 ; 7 9 : 6 8 0 6 8 5 .
© Copyright 2025 Paperzz





![[コラム随想]MEMSスケールでの材料破壊](http://s3.paperzz.com/store/data/005468567_1-bc59858eadaaf0d476f7b8ddf38c39ca-250x500.png)